Biomedicina
Analisi dell’architettura scheletrica di un modello animale in microgravità mediante Tomografia Assiale Computerizzata ad alta risoluzione
Alessandra Giuliani
Dipartimento di Scienze Cliniche Specialistiche ed Odontostomatologiche – Sezione Fisica
Facoltà di Medicina e Chirurgia, Università Politecnica delle Marche
L’osso è considerato uno dei tessuti più complessi del corpo umano a causa del continuo processo di rimodellamento che subisce in condizioni fisiologiche.
Ciò avviene non solo per sopportare i vincoli meccanici, ma anche per inibire il danno conseguente alla fatica meccanica. Inoltre, il rimodellamento è necessario per riparare le fratture, per mantenere vitali gli osteociti e per l’omeostasi del calcio.
Le cellule ossee agiscono sinergicamente, aumentando o diminuendo la massa ossea sulla base di diversi fattori: gli osteoblasti, le cellule che formano l’osso, regolano la deposizione delle molecole di matrice ossea, incluso il collagene di tipo I e una varietà di altre proteine non collaginee; gli osteoclasti, cellule giganti multinucleate, sono responsabili del riassorbimento della matrice ossea mineralizzata (Tavella et al., 2012).
Normalmente, il processo di rimodellamento consiste in una fase di costruzione lenta e in un riassorbimento più rapido, in un susseguirsi sinergico atto a mantenere una locomozione funzionale dello scheletro per contrastare la gravità terrestre. Pertanto, quando i movimenti e l’esercizio si riducono, come accade nelle persone allettate o negli astronauti durante i voli spaziali, lo scheletro non agisce più contro la gravità, con una rapida riduzione della massa e della densità ossea, causa principale di possibili fratture (Zhang et al., 2008 ).
L’architettura ossea e il suo rimodellamento vengono tradizionalmente studiati mediante radiografia a raggi X; purtroppo però questo metodo presenta alcune limitazioni, soprattutto dovute alla sovrapposizione di strutture anatomiche in singola proiezione che a sua volta può portare ad interpretazioni e diagnosi errate.
Negli anni ’70, lo sviluppo della prima apparecchiatura per tomografia computerizzata (TAC) ha rivoluzionato la diagnostica per immagini dei distretti ossei, dando nuovi stimoli per studi sempre più approfonditi.
La TAC è in grado di produrre ricostruzioni virtuali e tridimensionali (3D) dei tessuti di interesse, in modo non distruttivo e con un contrasto fino a mille volte migliore rispetto alle radiografie convenzionali (Claesson , 2001).
La microtomografia computerizzata (microCT) si basa sugli stessi principi fisici e metodologici della TAC convenzionale ma, mentre questa ha tipicamente una risoluzione lineare massima di circa 500 μm, alcuni strumenti per la microCT raggiungono una risoluzione spaziale fino a 200-300 nm (Cancedda et al., 2007), con un incremento di circa tre ordini di grandezza rispetto alla TAC.
Negli ultimi anni, la microCT è stata utilizzata in modo massiccio per ricostruire virtualmente, in modo non distruttivo e ad alta risoluzione, la complessa architettura del tessuto osseo. In effetti, nel campo della ricerca sulle ossa, sono stati sviluppati diversi metodi atti ad estrarre, dalle immagini di microCT, una serie di parametri quantitativi relativi alla micro-morfometria ossea. Ad esempio, il metodo Mean Intercept Length (MIL) è in grado di misurare lo spessore e la distanza media trabecolare in base ad alcune ipotesi geometrico-strutturali (Hildebrand & Ruegsegger, 1997a). È anche possibile bypassare queste ipotesi, estrapolando parametri che sono indipendenti da qualunque modello (Hildebrand & Ruegsegger, 1997b).
In questo scenario, la radiazione di sincrotrone (SR) si è dimostrata di fondamentale importanza negli studi di microCT a motivo delle sue caratteristiche peculiari, tra cui l’elevato rapporto segnale/rumore, l’elevato flusso di fotoni (che permette di ottenere dati ad alta risoluzione spaziale) e la possibilità di modulare l’energia dei fotoni, evitando effetti di beam-hardening.
La SR-microCT ha efficientemente supportato diversi studi sull’architettura ossea e sulla mappatura del livello di mineralizzazione di questa, a diversi livelli gerarchici (Lane et al., 2005; Nuzzo et al., 2002; Bousson et al., 2004), includendo anche l’imaging del sistema lacuno-canalicolare (Langer et al., 2012; Peyrin et al., 2014). Inoltre, la SR-microCT è stata anche impiegata per ricostruire, ad alta risoluzione, la complessa architettura del tessuto osseo in diverse condizioni genetiche e ambientali (Martin-Bardosa et al., 2003; Costa et al., 2013; Hesse et al., 2014), diventando negli anni uno strumento sempre più performante al fine di caratterizzare anche l’osso ingegnerizzato nei diversi distretti scheletrici (Cancedda et al., 2007, Giuliani et al., 2013; Giuliani et al., 2014; Mazzoni et al., 2017).
Inoltre, i recenti progressi nell’ambito della fisica e della tecnologia su cui si fonda la microCT hanno ulteriormente stimolato lo studio di siti scheletrici di carico mediante nuovi approcci di imaging. Questi ultimi non sono solo finalizzati all’ottenimento di immagini funzionali ma risultano anche di supporto allo studio di diverse malattie degenerative, spesso caratterizzate da perdita ossea in siti specifici. Un esempio notevole tra questi è quello derivante dalla permanenza in un ambiente senza gravità, come quello presente durante un volo spaziale.
Impatto della microgravità sull’osso
L’interesse per questo tipo di studio ha avuto inizio dopo le missioni Gemini, Apollo e Skylab, in seguito alle quali gli astronauti sperimentarono una forte demineralizzazione ossea accoppiata ad un aumento dell’escrezione di calcio. La conseguenza fu una procurata fragilità ossea, paragonabile a quella di soggetti costretti a letto per lunghi periodi di tempo (Wronski and Morey, 1983).
In effetti, la condizione dell’allettato è la più simile sperimentabile sulla Terra a quella subita dallo scheletro dell’astronauta dopo permanenza in gravità ridotta: viene infatti compromesso il rapporto tra formazione ossea e riassorbimento, con una conseguente perdita ossea accelerata da una maggiore attività osteoclastica (Donaldson et al., 1970; Leblanc et al., 1990, 1995).
Gli studi condotti sui cosmonauti a bordo della stazione spaziale russa MIR hanno confermato i risultati precedenti, evidenziando una perdita ossea significativa sulle ossa di carico (tibia) e massa ossea inalterata su quelle non portanti (radio) (Vico et al., 2000).
Un’analisi analoga sui membri dell’equipaggio, dopo 4-6 mesi di permanenza presso la Stazione Spaziale Internazionale (ISS), ha fornito numerosi dati morfometrici sull’osso corticale e trabecolare della colonna vertebrale e del sito d’anca, utilizzando la tomografia computerizzata quantitativa (QCT). Sono stati evidenziati sia l’assenza di perdita di osso nella colonna vertebrale che una perdita di mineralizzazione ossea corticale nell’anca, principalmente causata da un assottigliamento endocorticale (Lang et al., 2004).
Tuttavia, il punto debole degli studi precedentemente riportati era duplice: da un lato la ricerca soffriva del ridotto campione statistico, per il numero limitato di soggetti umani coinvolti nelle esperienze di volo, dall’altra i risultati sono stati sempre penalizzati in termini di affidabilità dal fatto che le strutture ossee osservate erano piccole rispetto alla limitata risoluzione spaziale dei dispositivi di imaging utilizzati (Lang et al., 2004).
Queste ragioni hanno condotto all’utilizzo crescente di modelli animali, che vanno dai ratti (Cosmi et al., 2009; Keune et al., 2015) ai pesci (Chatani et al., 2015), al fine di incrementare la dimensione statistica del campione (fornendo dati statistici più significativi) e per eseguire indagini di microCT sempre più informative, essendo quest’ultima una tecnica di imaging 3D ad alta risoluzione, in grado cioè di studiare strutture ossee di più piccole dimensioni rispetto alla TAC convenzionale.
Topi nello spazio
La maggior parte degli studi sulla microgravità è stata condotta utilizzando, come modello animale, il topo ed i suoi diversi strain. Questi esperimenti sono stati condotti nello spazio e, per ciascuna missione, sono state utilizzate gabbie sempre più sofisticate per l’alloggio degli animali.
In effetti, le condizioni ambientali possono influenzare la fisiologia e il comportamento dei topi sia sulla Terra che nello spazio. Blottner et al. (2009) analizzarono su 24 topi wild-type C57BL / 6JRj, ospitati per 25 giorni nel Prototipo di habitat MIS (Mice In Space), gli effetti del confinamento nella gabbia sul sistema muscolo-scheletrico. Il sistema MIS, completamente automatizzato, era una parte del dispositivo MSRM1 (Mouse Science Reference Module), prodotto da Alcatel Alenia Space Inc (Milano, Italia). Il Gruppo di Fisica del Dip. Di.S.C.O. dell’Università Politecnica delle Marche ha partecipato a questa ricerca tramite studi di SR-microCT su tre diversi siti ossei (calvaria, colonna vertebrale e femore) dei suddetti 24 topi. Confrontati con i topi alloggiati in gabbie convenzionali, i topi MIS non hanno rilevato cambiamenti significativi sia in termini di microarchitettura che con riferimento al grado di mineralizzazione in uno qualsiasi dei siti ossei studiati.
Tre missioni spaziali hanno fornito i primi dati documentati di microCT ad alta risoluzione in merito alle mutazioni scheletriche dei topi in volo: la missione shuttle STS-108, della durata di 12 giorni, con topi femmina C57BL6/J di 2 mesi, la missione shuttle STS-131 di 15 giorni, con topi femmina C57BL/6J di 16 settimane e la missione di 91 giorni a bordo della ISS con topi maschio di 2 mesi di tipo wild-type C57BL/10J (WT) e pleiotropina-transgenico (PTN-Tg).
Nel primo esperimento, Lloyd et al. (2015) hanno testato la capacità dell’Osteoprotegerin-Fc (OPG-Fc) di preservare la massa ossea durante il volo spaziale (SF). Ventiquattro ore prima del lancio, dodici topi per gruppo sono stati iniettati con OPG-Fc o mezzo inerte (VEH). I topi di controllo a terra (GC), di tipo VEH e OPG-Fc, sono stati allevati in condizioni ambientali che riproducevano quelle nello Space Shuttle, mentre i controlli (BL) sono stati sacrificati prima del lancio. Tutti i campioni ossei sono stati analizzati tramite un dispositivo di microCT da laboratorio (μCT20; Scanco Medical AG; Brüttisellen, Svizzera), al fine di studiare l’architettura trabecolare. Sono stati utilizzati i seguenti parametri sperimentali: dimensione del pixel di 9 μm, tensione di 55 KV, intensità di corrente di 145 mA e tempo di esposizione per proiezione (pp) di 200 ms. I siti di scansione erano la porzione trabecolare immediatamente distale rispetto alla cartilagine di accrescimento di tibia e omero. I parametri dell’osso trabecolare inclusi nello studio erano la frazione volumica di osso trabecolare (BV/TV), la densità di connettività (Conn.D), il numero di trabecole (Tb.N), la distanza media tra queste (Tb.Sp) e l’indice del modello di struttura (SMI). Nella tibia, la BV/TV di SF/VEH era inferiore del 26% rispetto ai GC/VEH, la Conn.D era inferiore del 27% (sebbene in modo non significativo), il Tb.Th era inferiore del 16% e lo SMI maggiore del 6%. Il volo spaziale non ha prodotto modifiche, come dedotto dall’analisi degli stessi parametri derivanti dai topi SF/OPG-Fc, quando confrontati con i GC/OPG-Fc. Nel sito dell’omero invece, sia il BV/TV che la Conn.D non sono stati modificati dal volo spaziale. In sintesi, questo studio di microCT ha dimostrato che un singolo trattamento con OPG-Fc prima del volo ha efficacemente impedito che si manifestassero gli effetti dannosi della microgravità sulle ossa del topo.
Nel secondo esperimento, Blaber et al. (2013) esposero otto topi a microgravità per testare se l’osteolisi osteocitica e l’arresto del ciclo cellulare durante l’osteogenesi potessero contribuire al riassorbimento osseo in condizioni di microgravità. Anche in questo caso è stato utilizzato un microtomografo da laboratorio (SkyScan 1174 scanner, Kontich, Belgio) per visualizzare e quantificare la morfometria ossea della regione dell’ischio destro. Le immagini sono state acquisite con i seguenti parametri sperimentali: dimensione del pixel di 6.77μm, tensione di scansione di 50 kV, 800mA di corrente e tempo di esposizione pp di 3,5 s. Queste analisi del bacino hanno mostrato che la microgravità aveva indotto una diminuzione del BV/TV del 6,29% e dello spessore dell’osso (B.Th) dell’11,91%, senza ridurre la densità di mineralizzazione ossea (BMD).
Successivamente, durante la missione italiana Mice Drawer System (MDS), sei topi sono stati esposti alla microgravità per 91 giorni presso la ISS (Cancedda et al., 2012) (Fig1). Questo volo spaziale è, ad oggi, il più lungo mai sperimentato: per questo motivo, ha fornito un vasta gamma di risultati, compresi numerosi studi sul transgene PTN, possibile protezione contro la perdita ossea dovuta alla assenza di gravità (Tavella et al., 2012).
Il Gruppo di Fisica del Dip. Di.S.C.O. dell’Università Politecnica delle Marche, all’interno di un ampio progetto di ricerca coordinato dal Prof. Ranieri Cancedda, ha perseguito la caratterizzazione tramite microCT dei reperti ossei, con risultati pubblicati su diverse riviste di impatto internazionale. L’imaging è stato eseguito presso la SYRMEP Beamline della ELETTRA Synchrotron Radiation Facility (Trieste, Italia), utilizzando una energia di 19 keV e una dimensione di pixel di 9μm. Le analisi si sono concentrate su due siti di carico, ovvero la porzione terza-inferiore del femore di sinistra (porzioni trabecolari e corticali) e il corpo vertebrale della VII lombare. Nelle ossa non portanti, l’analisi è stata circoscritta alla porzione sinistra dell’osso parietale (direzione trasversale dalla sutura sagittale al bordo). Le analisi di microCT hanno rivelato una perdita di massa ossea durante il volo spaziale in entrambi i ceppi WT e PTN, con una diminuzione del numero medio di trabecole (Tb.Nr) e un aumento della loro distanza media (Tb.Sp) dopo il volo (Fig2). Nello stesso studio è stato anche dimostrato che le ossa non portanti non sono influenzate dai vincoli di microgravità.

Figura 1. A. Il Progetto Mice Drawer System (MDS). B. L’astronauta Nicole Stott, ingegnere aerospaziale della missione 20/21, vicino alla facility MDS nel laboratorio KIBO della ISS (credits NASA). C. Il modello di volo Mice Drawer System nel dettaglio. https://doi.org/10.1371/journal.pone.0032243.g001
Un’altra missione, la Bion-M1, ha offerto una nuova opportunità per caratterizzare, mediante microCT, i cambiamenti scheletrici di topi maschi C57/BL6 adulti (23 settimane di età al lancio) dopo 30 giorni di voli spaziali e un periodo di recupero di 8 giorni (Gerbaix et al., 2017).
Nel protocollo sperimentale di quest’ultima missione erano inclusi due gruppi di controllo a terra come per la missione MDS: un gruppo Habitat Control, allevato in gabbie analoghe a quelle nello spazio, e un gruppo Control, allevato in gabbie standard. Tutti i femori e le vertebre L3 e T12 (5/6 animali per gruppo) sono stati scansionati con un dispositivo di tomografia ad alta risoluzione (VivaCT40, Scanco Medical, Bassersdorf, Svizzera), utilizzando pixel di 12,5 μm e un protocollo di analisi precedentemente adottato e descritto da (David et al., 2003). Tramite confronto con il controllo a terra, si è scoperto che il confinamento nella gabbia utilizzata in volo influenza negativamente la morfometria del femore e delle vertebre lombari (ma non quella delle vertebre toraciche): infatti, le vertebre L3 e le trabecolari del femore presentavano un rapporto BV/TV e una Conn.D ridotte nel Controllo Habitat rispetto al gruppo di controllo. Inoltre, il volume specifico trabecolare BV/TV delle vertebre L3 risultava ridotto nel gruppo di volo rispetto ad entrambi i gruppi di controllo (-35,7 % rispetto all’Habitat Control e -56,5% rispetto ai Control; p <0,0033), molto probabilmente a causa della diminuzione di Tb.N e Tb.Th. Una riduzione di massa ossea ancora più rilevante è stata riscontrata nei femori del gruppo di volo rispetto ai gruppi di controllo (-85,2% rispetto ai Control, p <0,0003; -64,8% rispetto all’Habitat Control, p <0,017).
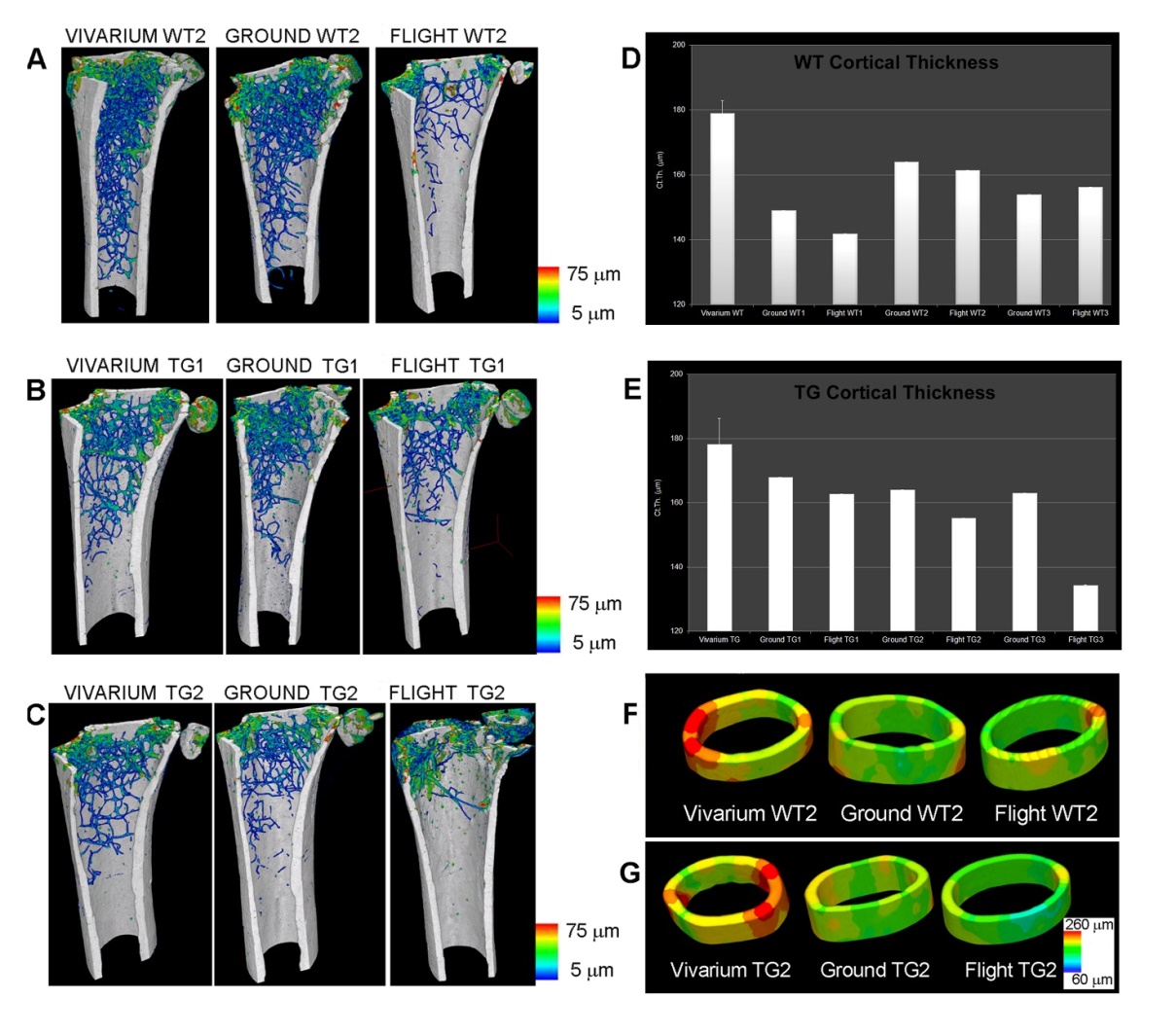
Figura 2. Missione spaziale MDS. Femori di topo alloggiati per 3 mesi nella Stazione Spaziale Internazionale (ISS). (A-C) Wt2 (A), PTN-Tg1 (B), PTN-Tg2 (C): mappa a colori della distribuzione di spessore trabecolare di un controllo (Vivarium) (campione rappresentativo), dei topi allevati a terra in gabbie MDS (Ground) e di quelli in volo (Flight). (D, E) Quantificazione della distribuzione dello spessore corticale in topi Wt (D) e PTN-Tg (E). Valore del Vivarium: media su 3 topi di controllo. (F, G) Mappe a colori di ricostruzioni 3D rappresentative dello spessore corticale nel Vivarium, Ground e Flight Wt2 (F) e nel Vivarium, Ground e Flight PTN-Tg2 (G). (A-C) sono stati originariamente pubblicati nella Figura 1 di (Tavella et al., 2012), (D-G) nella Figura 3 di (Tavella et al., 2012). Dati riproposti nella Fig.1 di Giuliani et al. (2018).
Parametri trabecolari simili (Fig3) sono stati trovati nel gruppi post-volo (Flight) e in quello che dopo il volo aveva sperimentato la permanenza per 8 giorni in un contesto di gravità ripristinata (Flight+Rec), deducendo che 8 giorni costituiscono un intervallo temporale non sufficiente per iniziare il recupero della struttura ossea originaria.
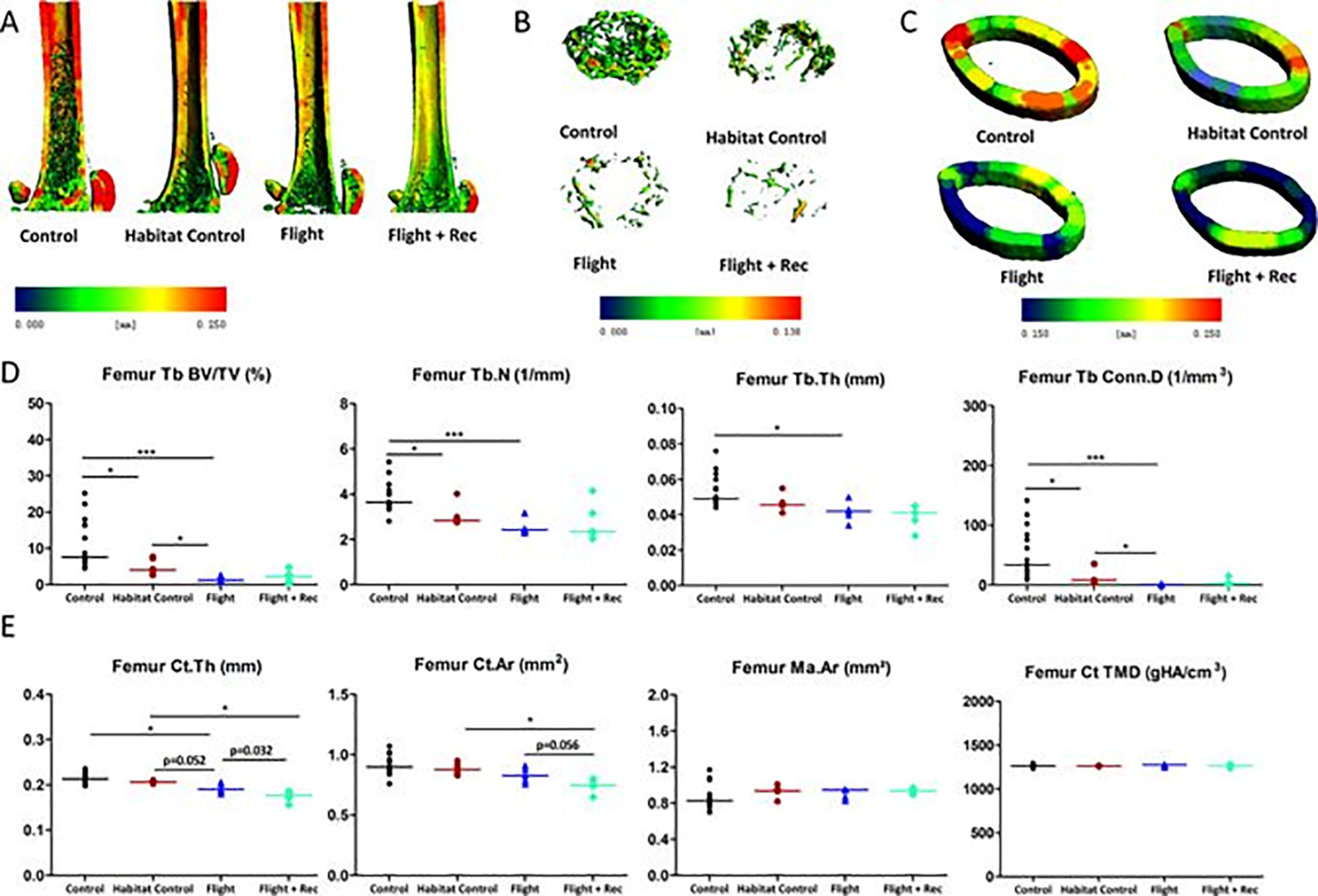
Figura 3. Effetto delle condizioni di alloggiamento nel veicolo spaziale, del volo spaziale e del recupero dopo atterraggio. Da Gerbaix et al., 2017. (A) Microarchitettura del femore; (B) microarchitettura della metafisi trabecolare (Tb) e (C) osso corticale (Ct) al centro della diafisi. Una mappa cromatica proporzionale agli spessori ossei illustra la diminuzione della massa ossea, del volume trabecolare (Tb) e dello spessore corticale (Ct) indotti dalla microgravità e a seguito di recupero a terra per 8 giorni. (D) Parametri di microarchitettura trabecolare e (E) corticale. Per la frazione trabecolare: volume osseo specifico (BV/TV), numero trabecolare (Tb.N), spessore trabecolare (Tb.Th) e densità di connettività (Conn.D); per la frazione corticale: spessore (Ct.Th), area (Ct.Ar), densità di mineralizzazione del tessuto (TMD) e area del midollo (Ma.Ar). * p <0,017, ** p <0,003
Nello stesso studio (Gerbaix et al., 2017), sono state studiate tramite microCT cinque sezioni corticali del femore per gruppo. Le analisi sono state condotte alla beamline ID19 della European Synchrotron Radiation Facility (Grenoble, Francia), con una dimensione di pixel di 0,7 μm, 2000 proiezioni su 360° di rotazione del campione e un’energia del fascio fotonico pari a 26 keV. L’analisi 3D si è focalizzata sulle diverse migliaia di lacune osteocitarie, ottenendo dati sul volume totale lacunare (Lc.V, mm3), sulla densità delle lacune (N.Lc/TV) e sulla densità del volume lacunare Lc.V/TV (%). Essendo la forma di una lacuna osteocitaria approssimabile ad un ellissoide, gli autori hanno utilizzato i momenti del secondo ordine per misurare in modo efficiente le lunghezze degli assi principali dell’ellissoide di best-fitting. Questa analisi estremamente sofisticata ha mostrato che, nel gruppo Flight, le lacune degli osteociti avevano un volume più piccolo e con una forma più sferica. Inoltre, il numero di lacune vuote risultava essere aumentato in modo significativo (+ 344%) rispetto al gruppo Habitat Control. Questi dati hanno dimostrato che la microgravità può causare la morte degli osteociti, a sua volta responsabile del riassorbimento osseo con conseguente perdita di massa. Più recentemente lo stesso gruppo di ricerca (Gerbaix et al., 2018), studiando tramite microCT le ossa della caviglia (calcagno, navicolare e talus), ha osservato che la perdita ossea indotta dal volo è specifica per sito e compartimento, mentre la densità di mineralizzazione del tessuto osseo rimane preservata. Inoltre, otto giorni dopo l’atterraggio, lo stato delle ossa della caviglia sembra addirittura peggiorato rispetto alla situazione immediatamente dopo l’atterraggio.
La missione Bion-M1 di 30 giorni ha anche offerto la possibilità di studiare sei topi maschi C57BL/6 (19-20 settimane di vita al momento del lancio). Gli animali furono sacrificati 13-15 ore dopo l’atterraggio. Otto topi di controllo sono stati allevati a terra durante lo stesso periodo di 30 giorni, in gabbie standard del vivarium. Segmenti delle vertebre caudali dei topi Flight e di controllo sono stati sottoposti a carico fino a rottura (Berg-Johansen et al., 2016). Dopo questi test, gli stessi segmenti sono stati analizzati tramite microCT (μCT 50, SCANCO Medical, Brüttisellen, Svizzera) per quantificare la microarchitettura trabecolare e la BMD. I campioni sono stati scansionati utilizzando una dimensione di pixel di 4 μm, tensione di 55 kV e intensità di corrente a 109 μA. Si è osservato che il volo spaziale aveva ridotto in modo significativo il rapporto BV/TV, la BMD e la Tb.Th vertebrale, fornendo una possibile spiegazione della tendenza dei campioni di volo ad andare incontro a rottura proprio nell’osso epifisario.
Questi risultati di microCT, combinati con quelli dei test di flessione meccanica, indicano che la perdita ossea vertebrale durante il volo spaziale può compromettere la capacità di flessione della colonna vertebrale, contribuendo negli astronauti ad aumentare il rischio di ernia del disco.
Nonostante le informazioni importanti acquisite grazie alle due missioni precedentemente descritte (la MDS di 91 giorni e la Bion-M1 di 30 giorni), i protocolli sperimentali di queste sembravano inizialmente carenti nella gestione dei topi di controllo. I controlli infatti, trovandosi a terra invece che nello spazio, potrebbero aver influenzato i risultati falsandoli. Infatti questa scelta non tiene in considerazione alcuni fattori, come la radiazione cosmica, l’ambiente microbico, la vibrazione di volo, lo shock e l’accelerazione semi-costante durante le fasi di lancio e di ritorno all’interno del veicolo spaziale.
Di conseguenza, Shiba et al. (2017) hanno recentemente sviluppato una nuova piattaforma sperimentale per generare gravità artificiale nello spazio.
Questo studio è stato condotto nell’ambito di un progetto della Japan Aerospace Exploration Agency (JAXA) e si è incentrato sullo studio dell’impatto della gravità parziale (partial g) e della microgravità (μg) sui topi, utilizzando gabbie (habitat cage units – HCU) di nuova concezione, cioè equipaggiate con centrifuga per esperimenti biologici nella ISS.
Nella prima missione del progetto, 12 topi maschi C57BL/6J (5 settimane di vita al lancio) sono stati sottoposti a μg oppure a gravità artificiale pari a quella della Terra (1g) (Fig4). I topi sottoposti a μg galleggiavano all’interno dell’HCU, mentre i topi sottoposti a 1g artificiale poggiavano regolarmente sul fondo dell’HCU, con una centrifugazione a 77 giri/min (Fig5). Dopo 35 giorni, tutti i topi sono stati riportati sulla Terra e studiati tramite microCT. Il femore destro di ogni topo è stato analizzato utilizzando uno scanner ScanXmate-A100S (Comscantechno, Yokohama, Giappone).
La microCT ha rivelato che il carico di gravità artificiale riduceva la perdita ossea indotta dalle condizioni di microgravità. Infatti, diminuzioni significative della densità ossea erano visibili nel femore dei topi μg, mentre i topi sottoposti a 1g artificiale mantenevano la stessa densità ossea dei topi di controllo a terra. Questi dati hanno quindi ulteriormente confermato che la perdita ossea degli esperimenti precedenti, la MDS di 91 giorni e la Bion-M1 di 30 giorni, era specificamente dovuta alla permanenza in microgravità.

Figura 4. Gabbie per topi di nuova concezione per l’esperimento spaziale. Da Shiba et al., 2017. (a) Habitat Cage Unit (HCU). L’HCU ospita un topo per ogni gabbia. 1: Serbatoio dell’acqua, 2: Fotocamera / LED, 3: Sensore di temperatura, 4: Ingresso acqua per pulizia, 5: Ugelli acqua, 6: Tergicristallo per fotocamera, 7: Cartuccia per alimenti, 8: Pavimento / pareti in policarbonato. All’interno dell’HCU è alloggiata una fornitura di cibo e acqua per 7 giorni. (b) Gabbia di trasporto (TCU). La TCU contiene 12 gabbie cilindriche per unità. Ogni gabbia cilindrica ospita un topo. La TCU supporta fino a 10 giorni di cibo e acqua durante le fasi di lancio e atterraggio. 1: area abitativa con cibo, 2: serbatoio dell’acqua, 3: cibo, 4: ugello acqua, 5: pavimento / pareti in policarbonato. (c) Attrezzatura per esperimenti biologici dotata di centrifuga (CBEF). Il CBEF ha due comparti: la sezione micro-g (MG) e la sezione artificial-g (AG) dotata di centrifuga. (d) sezione AG. La centrifuga ospita sei HCU e il raggio di rotazione in cui entra in contatto con il pavimento è di 0,15 m.
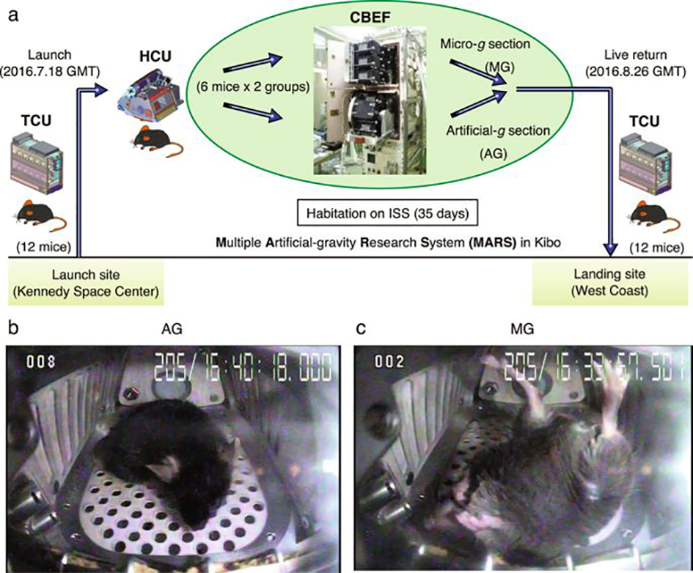
Figura 5. Schema concettuale del primo Progetto-Mouse della JAXA. Da Shiba et al., 2017. (a) Dodici topi maschi (C57BL/6J) sono stati lanciati dallo SpX9 dal Kennedy Space Center il 18 luglio 2016. Sulla ISS, i topi sono stati divisi in due gruppi (sei topi in MG e sei topi in AG). Dopo 35 giorni di permanenza sulla ISS, i topi della TCU sono stati collocati nella capsula Dragon e poi fatti ammarare nell’Oceano Pacifico vicino alla West Coast, il 26 agosto 2016. I topi sono stati trasportati al laboratorio per l’osservazione comportamentale prima e la dissezione due giorni dopo. Immagini rappresentative dell’alloggiamento a bordo per un topo AG (b) e un topo MG (c).
I nuovi studi di frontiera
La maggior parte dei dati derivanti dagli studi effettuati sui topi sono risultati in accordo con le osservazioni precedentemente raccolte sull’uomo, confermando l’utilità del modello murino nelle indagini in merito agli effetti dei voli spaziali sulla microarchitettura scheletrica. Nonostante ciò, i risultati ottenuti andrebbero analizzati con cautela e attenzione, considerando le condizioni sperimentali specifiche di ogni singolo esperimento, come età dell’animale, sesso, peso corporeo, e altre variabili; queste infatti potrebbero influenzare il risultato dello studio (Tavella et al., 2012). Questo fatto costituisce in sé un incentivo importante per continuare la ricerca sui diversi modelli animale sottoposti all’esperienza di volo spaziale, da un lato standardizzando i parametri dell’indagine, dall’altro approfondendo gli studi della ultrastruttura ossea mediante nanotomografia, possibilmente basata sulla radiazione di sincrotrone (Langer et al., 2012; Pacureanu et al., 2012; Hesse et al., 2015), come già effettuato in via esplorativa da Gerbaix et al. (2017) dopo la missione Bion-M1 di 30 giorni.
Inoltre, la possibilità di simulare un carico artificiale sul sistema muscolo-scheletrico, come nelle condizioni di ipergravità (Canciani et al., 2015) rappresenta un indubbio vantaggio per la ricerca scientifica in questo ambito, poiché può consentire ai ricercatori di evitare la complessità degli studi di microgravità reale e rende possibile l’analisi di un numero maggiore di soggetti, migliorando così il livello di affidabilità di tali studi.
Bibliografia
- Berg-Johansen, B., Liebenberg, E.C., Li, A., Macias, B.R., Hargens, A.R., and Lotz, J.C. (2016). Spaceflight-induced bone loss alters failure mode and reduces bending strength in murine spinal segments. J Orthop Res. 34(1):48-57. doi: 10.1002/jor.23029
- Blaber, E.A., Dvorochkin, N., Lee, C., Alwood, J.S., Yousuf, R., Pianetta, P., et al. (2013). Microgravity induces pelvic bone loss through osteoclastic activity, osteocytic osteolysis, and osteoblastic cell cycle inhibition by CDKN1a/p21. PLoS One. 18;8(4):e61372. doi: 10.1371/journal.pone.0061372
- Blottner, D., Serradj, N., Salanova, M., Touma, C., Palme, R., Silva, M., et al. (2009). Morphological, physiological and behavioural evaluation of a ‘Mice in Space’ housing system. J Comp Physiol B. 179(4):519-33. doi: 10.1007/s00360-008-0330-4
- Bousson, V., Peyrin, F., Bergot, C., Hausard, M., Sautet, A., and Laredo, J.D. (2004). Cortical Bone in the Human Femoral Neck: Three-Dimensional Appearance and Porosity Using Synchrotron Radiation. J Bone Miner Res 19:794–801
- Cancedda, R., Cedola, A., Giuliani, A., Komlev, V., Lagomarsino, S., Mastrogiacomo, M., et al. (2007). Bulk and interface investigations of scaffolds and tissue-engineered bones by X-ray microtomography and X-ray microdiffraction. Biomaterials 28(15), 2505-2524
- Cancedda, R., Liu, Y., Ruggiu, A., Tavella, S., Biticchi, R., Santucci, D., et al. (2012). The Mice Drawer System (MDS) experiment and the space endurance record-breaking mice. PLoS One. 7(5):e32243
- Canciani, B., Ruggiu, A., Giuliani, A., Panetta, D., Marozzi, K., Tripodi, M., et al. (2015). Effects of long time exposure to simulated micro- and hypergravity on skeletal architecture. Journal of the Mechanical Behavior of Biomedical Materials 51: 1–12
- Chatani, M., Mantoku, A., Takeyama, K., Abduweli, D., Sugamori, Y., Aoki, K., et al. (2015). Microgravity promotes osteoclast activity in medaka fish reared at the international space station. Scientific Reports 5, Article number: 14172. doi:10.1038/srep14172
- Claesson, T. (2001). A medical imaging demonstrator of computed tomography and bone mineral densitometry. Stockholm: Universitetsservice US AB; 2001
- Cosmi, F., Steimberg, N., Dreossi, D., and Mazzoleni, G. (2009). Structural analysis of rat bone explants kept in vitro in simulated microgravity conditions. J Mech Behav Biomed Mater. 2(2):164-72. doi: 10.1016/j.jmbbm.2008.06.004
- Costa, D., Lazzarini, E., Canciani, B., Giuliani, A., Spanò, R., Marozzi, K., et al. (2013). Altered bone development and turnover in transgenic mice over-expressing lipocalin-2 in bone. J Cell Physiol. 228(11):2210-21. doi: 10.1002/jcp.24391
- David, V., Laroche, N., Boudignon, B., Lafage-Proust, M.H., Alexandre, C., Ruegsegger, P., et al. (2003). Noninvasive in vivo monitoring of bone architecture alterations in hindlimb-unloaded female rats using novel three-dimensional microcomputed tomography. J Bone Miner Res. 18(9):1622-31
- Donaldson, C.L., Hulley, S.B., Vogel, J.M., Hattner, R.S., Bayers, J.H., and McMillan, D,E. (1970). Effect of prolonged bed rest on bone mineral. Metabolism 19: 1071–1084
- Gerbaix, M., Gnyubkin, V., Farlay, D., Olivier, C., Ammann, P., Courbon, G., Laroche, N., Genthial, R., Follet, H., Peyrin, F., Shenkman, B.S., Gauquelin-Koch, G., & Vico, L. (2017). Erratum: One-month spaceflight compromises the bone microstructure, tissue-level mechanical properties, osteocyte survival and lacunae volume in mature mice skeletons. Scientific Reports 7(1):2659. doi: 10.1038/s41598-017-03014-2
- Gerbaix M, White H, Courbon G, Shenkman B, Gauquelin-Koch G and Vico L (2018) Eight Days of Earth Reambulation Worsen Bone Loss Induced by 1-Month Spaceflight in the Major Weight-Bearing Ankle Bones of Mature Mice. Front. Physiol. 9:746. doi: 10.3389/fphys.2018.00746
- Giuliani, A., Manescu, A., Langer, M., Rustichelli, F,, Desiderio, V., Paino, et al. (2013). Three years after transplants in human mandibles, histological and in-line HT revealed that stem cells regenerated a compact rather than a spongy bone: biological and clinical implications. Stem Cells Translational Medicine 2, 316-324
- Giuliani, A., Manescu, A., Larsson, E., Tromba, G., Luongo, G., Piattelli, A., et al. (2014). In Vivo Regenerative Properties of Coralline-Derived (Biocoral) Scaffold Grafts in Human Maxillary Defects: Demonstrative and Comparative Study with Beta-Tricalcium Phosphate and Biphasic Calcium Phosphate by Synchrotron Radiation X-Ray Microtomography. Clin Implant Dent Relat Res.16(5), 736-50
- Giuliani, A., Mazzoni, S., Ruggiu, A., Canciani, B., Cancedda, R., & Tavella, S. (2018). High-Resolution X-Ray Tomography: A 3D Exploration Into the Skeletal Architecture in Mouse Models Submitted to Microgravity Constraints. Frontiers in Physiology, 9, 181. http://doi.org/10.3389/fphys.2018.00181
- Hesse, B., Langer, M., Varga, P., Pacureanu, A., Dong, P., Schrof, S., et al. (2014). Alterations of Mass Density and 3D Osteocyte Lacunar Properties in Bisphosphonate-Related Osteonecrotic Human Jaw Bone, a Synchrotron µCT Study. PLoS ONE, 9(2), e88481. http://doi.org/10.1371/journal.pone.0088481
- Hesse, B., Varga, P., Langer, M., Pacureanu, A., Schrof ,S., Männicke, N., et al. (2015). Canalicular network morphology is the major determinant of the spatial distribution of mass density in human bone tissue: evidence by means of synchrotron radiation phase-contrast nano-CT. J Bone Miner Res. 30(2):346-56. doi: 10.1002/jbmr.2324
- Hildebrand, T., and Ruegsegger, P. (1997a). Quantification of bone microarchitecture with the Structure Model Index. Comput Meth Biomech Biomed Eng 1:15–23
- Hildebrand, T., and Ruegsegger, P. (1997b). A new method for the model independent assessment of thickness in three-dimensional images. J Microsc 185:67–75
- Jia, F., Françoise, P., Malaval, L., Laurence, V. and Marie-Hélène, L.P. (2010), Imaging and Quantitative Assessment of Long Bone Vascularization in the Adult Rat Using Microcomputed Tomography. Anat Rec, 293: 215–224. doi:10.1002/ar.21054
- Keune, J.A., Branscum, A.J., Iwaniec, U.T., and Turner, R.T. (2015). Effects of Spaceflight on Bone Microarchitecture in the Axial and Appendicular Skeleton in Growing Ovariectomized Rats. Sci Rep. 22;5:18671. doi: 10.1038/srep18671
- Lane, N.E., Yao, W., Balooch, M., Nalla, R.K., Balooch, G., Habelitz, S., et al. (2005). Glucocorticoid-Treated Mice Have Localized Changes in Trabecular Bone Material Properties and Osteocyte Lacunar Size That Are Not Observed in Placebo-Treated or Estrogen-Deficient Mice. J Bone Miner Res 21:466–476
- Lang, T., LeBlanc, A., Evans, H., Lu, Y., Genant, H., and Yu, A. (2004). Cortical and trabecular bone mineral loss from the spine and hip in long-duration spaceflight. J Bone Miner Res 19: 1006–1012
- Langer, M., Pacureanu, A., Suhonen, H., Grimal, Q., Cloetens, P., and Peyrin, F. (2012). X-Ray Phase Nanotomography Resolves the 3D Human Bone Ultrastructure. PLoS ONE 7(8), e35691
- Leblanc, A.D., Schneider, V.S., Evans, H.J., Engelbretson, D.A., and Krebs, J.M. (1990). Bone mineral loss and recovery after 17 weeks of bed rest. J Bone Miner Res 5: 843–850
- LeBlanc, A., Schneider, V., Spector, E., Evans, H., Rowe, R., Lane, H., et al. (1995). Calcium absorption, endogenous excretion, and endocrine changes during and after longterm bed rest. Bone 16: 301S–304S
- Lloyd, S.A, Morony, S.E., Ferguson, V.L., Simske, S.J., Stodieck, L.S., Warmington, K.S., et al. (2015). Osteoprotegerin is an effective countermeasure for spaceflight-induced bone loss in mice. Bone. 81:562-72. doi: 10.1016/j.bone.2015.08.021
- Mazzoni, S., Mohammadi, S., Tromba, G., Diomede, F., Piattelli, A., Trubiani O, et al. (2017). Role of cortico-cancellous heterologous bone in human periodontal ligament stem cell xeno-free culture studied by Synchrotron radiation phase-contrast microtomography. Int. J. Mol. Sci. 18(2), 364
- Martín-Badosa, E., Amblard, D., Nuzzo, S., Elmoutaouakkil, A., Vico, L., and Peyrin, F. (2003). Excised bone structures in mice: imaging at three-dimensional synchrotron radiation micro CT. Radiology. 229(3):921-8
- Nuzzo , S., Lafage-Proust, M.H., Martin-Badosa, E., Boivin, G., Thomas, T., Alexandre, C., et al. (2002). Synchrotron Radiation Microtomography Allows the Analysis of Three-Dimensional Microarchitecture and Degree of Mineralization of Human Iliac Crest Biopsy Specimens: Effects of Etidronate Treatment. J Bone Miner Res 17:1372–1382
- Pacureanu, A., Langer, M., Boller, E., Tafforeau, P., Peyrin, F. (2012). Nanoscale imaging of the bone cell network with synchrotron X-ray tomography: optimization of acquisition setup. Med Phys. 39(4):2229-38. doi: 10.1118/1.3697525
- Peyrin, F., Dong, P., Pacureanu, A., and Langer, M. (2014). Micro- and nano-CT for the study of bone ultrastructure. Curr Osteoporos Rep.12(4), 465-74
- Shiba, D., Mizuno, H., Yumoto, A., Shimomura, M., Kobayashi, H., Morita, H., et al. (2017). Development of new experimental platform ‘MARS’-Multiple Artificial-gravity Research System-to elucidate the impacts of micro/partial gravity on mice. Sci Rep. 7(1):10837. doi: 10.1038/s41598-017-10998-4
- Tavella, S., Ruggiu, A., Giuliani, A., Brun, F., Canciani, B., Manescu, A., et al. (2012). Bone turnover in wild type and pleiotrophin-transgenic mice housed for three months in the International Space Station (ISS). PLoS One 7(3), e33179.Vico, L., Collet, P.,
- Guignandon, A., Lafage-Proust, M.H., Thomas, T., Rehaillia, M., et al. (2000). Effects of long-term microgravity exposure on cancellous and cortical weight-bearing bones of cosmonauts. Lancet. 6;355(9215):1607-11
- Wronski, T.J., and Morey, E.R. (1983). Alterations in calcium homeostasis and bone during actual and simulated space flight. Med Sci Sports Exerc 15: 410–414
- Zhang, P., Hamamura, K., and Yokota, H. (2008). A brief review of bone adaptation to Unloading. Genomics Proteomics Bioinformatics 6: 4–7